
Mécanismes physiques de la morphogenèse de réseaux biologiques
Participants : Annemiek Cornelissen, Marc Durand, Giulio Facchini
Les réseaux de transport biologiques sont essentiels au fonctionnement de nombreux organismes. On peut citer le système vasculaire des vertébrés (de l’aorte au lit capillaire) et des plantes (des racines à la nervation des feuilles), le système bronchique et le mycélium des colonies fongiques. Par ailleurs, d’autres réseaux poreux fait de minérale ou de fibres organiques constituent la structure interne des endosquelettes de multiples animaux et façonnent leur propriétés mécaniques (échinodermes) ou même optiques (lépidoptères). Ces structures auto-organisées présentent un certain nombre de propriétés (efficacité, adaptabilité, résilience) qui sont hautement souhaitables pour des applications techniques. La compréhension de leur formation et de leur évolution a également des applications médicales évidentes, car de nombreuses maladies sont associées à des évolutions pathologiques de leur structure. structure. Il est bien documenté que la croissance et le remodelage des réseaux de transport biologiques sont influencés par des facteurs mécaniques. A l’aide d’organismes modèles, le myxomycète Physarum polycephalum et la méduse Aurelia aurita, nous étudions l’importance des contraintes mécaniques (émanant des tissus environnants ou des flux internes) sur la formation et le remodelage des réseaux biologiques. Nous nous intéressons également aux mécanismes d’auto-organisation de l’activité contractile, responsables des flux générés dans ces organismes dépourvus de cœur. À l’aide de l’oursin Paracentrotus lividus, nous étudions la fabrication d’un réseau biominéral poreux par le tissu squelettogenique qui occupe ses pores. Nous nous intéressons particulièrement au rôle du cytosquelette dans ce processus de croissance.
Morphogenèse du réseau de canaux du système gastrovasculaire de la méduse Aurelia aurita
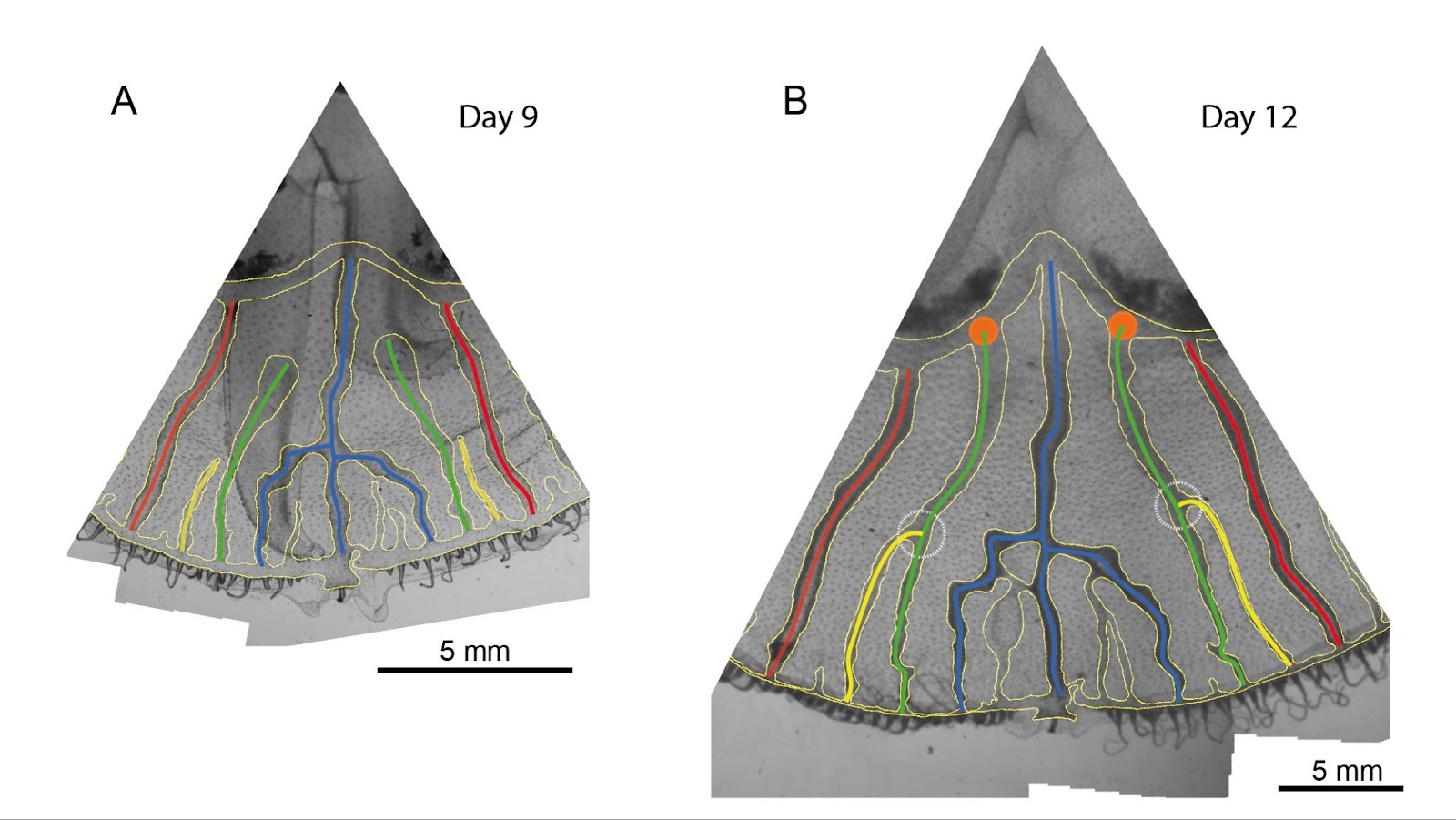
Nous étudions la morphogenèse du réseau de canaux du système gastrovasculaire de la méduse Aurelia aurita. Nous cherchons à caractériser le réseau à une échelle macroscopique. En quantifiant la croissance par des observations quotidiennes du réseau, nous avons développé des algorithmes qui nous permettent de retracer la croissance du réseau de canaux. En utilisant l’historique de croissance, nous sommes en mesure d’évaluer quantitativement la diversité des motifs. Nous supposons que les motifs proviennent d’instabilités auto-organisées. La première instabilité que nous observons est en analogie avec la propagation des fissures. Les muscles de la sous-ombrelle se contractent, générant ainsi une compression dans le feuillet endodermique dans laquelle les canaux se développent. Comme les fissures, les canaux creux et mous libèrent la contrainte de compression dans le feuillet endodermique. Pour comprendre comment la contrainte de compression est distribuée dans l’endoderme, nous avons observé de petites déformations dues aux contractions musculaires lorsque la méduse est couchée à plat sous le microscope et de grandes déformations pendant la nage. Des modèles simulations nous ont permis d’estimer la distribution des contraintes dans le feuillet endodermique autour des canaux. La position de la contrainte maximale est en corrélation avec la croissance biaisée des bourgeons que nous observons chez les méduses. La deuxième instabilité est en analogie avec les instabilités de branchement des systèmes hydrodynamiques, comme l’expérience bien connue de Saffman-Taylor, injectant de l’air dans de l’huile. Lorsque la mobilité de la phase envahissante (air) est plus grande que la mobilité de la phase déplacée (huile), la frontière entre les phases devient instable et le front se brise en doigts. Lorsque le rapport des mobilités des deux phases est faible, ces doigts peuvent se reconnecter. Un cas extrême se produit lorsqu’un doigt se brise et que le doigt voisin se connecte toujours. C’est exactement cette reconnexion corrélée au temps que nous observons dans le réseau de canaux d’Aurélia.
Activité contractile et développement du réseau tubulaire de P. polycephalum
Dans sa phase plasmode, P. polycephalum est un organisme unicellulaire géant qui développe alors un réseau tubulaire dans lequel des écoulements sont générés par la contraction active de la couche membraneuse entourant les “veines”, suppléant ainsi aux écoulements diffusifs trop lents. Toutefois, la génération d’écoulements sur de grandes distances nécessite la coordination de ces contractions qui prend la forme d’ondes de contraction. Tout au long de sa vie, l’organisme est capable de remodeler l’architecture de son réseau via un contrôle local de la transition sol-gel. Afin d’élucider les mécanismes de couplage entre activité contractile, flux, et formation (ou remodelage) du réseau, nous avons étudié la coordination de l’activité contractile et le remodelage du réseau dans des spécimens confinées dans des chambres de géométrie annulaire dont on peut faire varier le rayon et la largeur (cf. figure ci-contre). Cette géométrie permet de supprimer tout axe antéro-postérieur prédéfini de l’organisme, et bloque la croissance ou la migration lorsque la chambre est entièrement remplie. Cette géométrie rend également le système unidimensionnel. Nous avons construit un dispositif original d’imagerie en transmission placé dans une enceinte à humidité et température contrôlées. Nous avons également développé des outils spécifiques de traitement d’images et d’analyse du signal afin de caractériser entièrement la structure du réseau (topologie, longueurs et diamètres des veines, conductances) – avec une résolution inédite permettant d’identifier plusieurs milliers de veines – ainsi que les ondes de contraction (phase instantanée, amplitude, longueur d’onde, fréquence, vitesse de phase) et les débits traversant le réseau. Ce montage nous a permis de caractériser les différents modes contractiles stables qui pouvaient s’établir dans la chambre annulaire et leur durée de vie respective. Ces résultats ont permis de discriminer les différents modèles de couplage entre contractions et écoulements existants dans la littérature. Nous avons également mis en évidence et quantifié la réorientation aux temps longs des veines du réseau selon la direction orthoradiale

Morphogenèse de la microsctructure du squelette de P. lividus
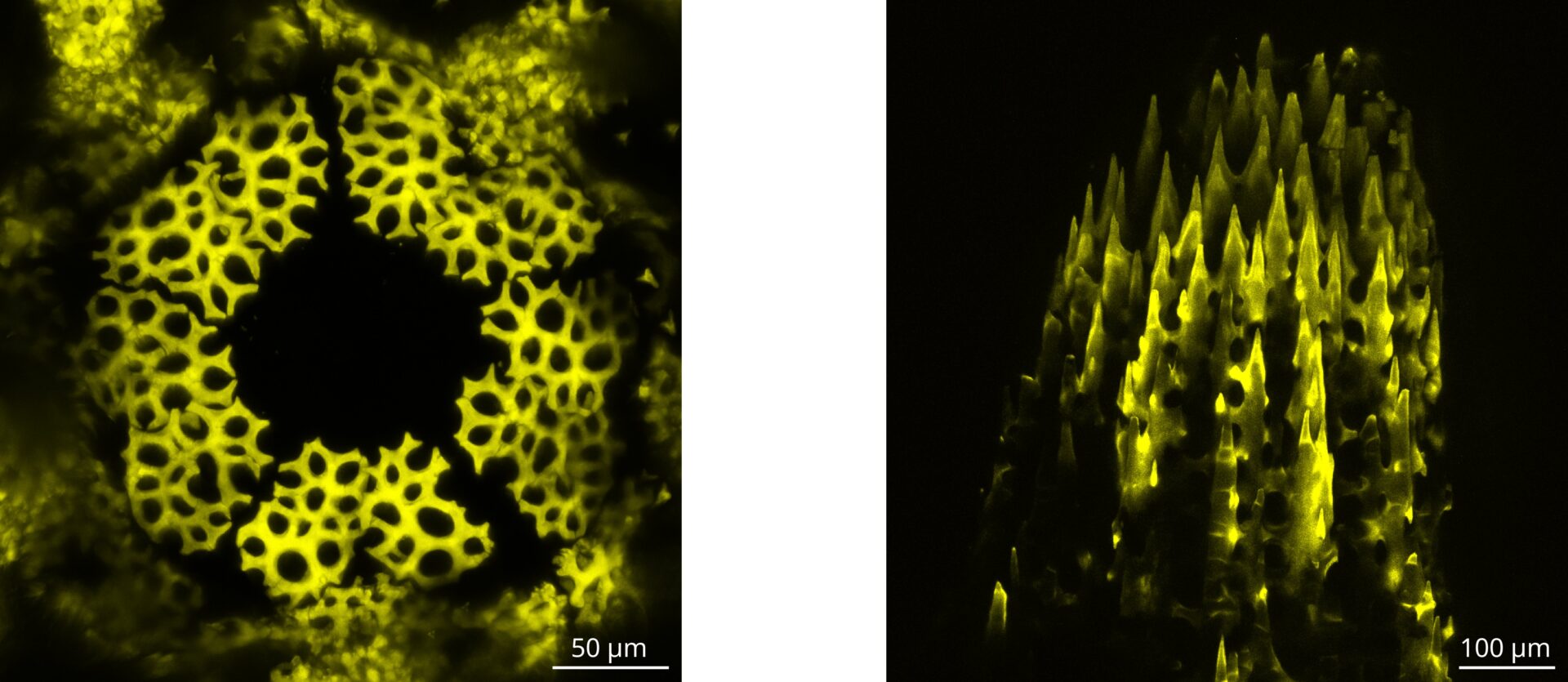
Les échinodermes, comme les oursins, les étoiles de mer ou les concombres de mer construisent un squelette biominéral en calcite, avec une microstructure complexe qui est un exemple formidable d’auto-organisation et prend le nom de stéréome. Ceci est un réseau microporeux 3D avec une surface courbée en forme de selle, qui ressemble à des surfaces minimales et qui lui donne des propriétés mécaniques optimales. Ce genre de géométries ont intrigué les scientifiques pendant des siècles, car elles peuvent émerger de manière spontanée comme résultat de la minimisation d’une énergie d’interface comme la tension de surface. Dans le cas du stéréome, nous savons qu’il se forme par la biominéralisation de milliers de trabécules micrométriques qui croissent par les pointes, font des branchements et fusionnent entre elles. On sait aussi que la croissance du squelette bio minéral a lieu par l’ajout progressif de petites particules de calcite amorphes qui sont synthétisées par les cellules qui habitent le tissu remplissant les pores interconnectés du stéréome. Cependant, le mécanisme général qui permet aux cellules de diriger la déposition de nouveau minéral reste inconnu.
Nous étudions la croissance du stéréome chez l’oursin Paracentrotus lividus, et en particulier nous cherchons à comprendre l’origine de sa courbure mais aussi le rôle joué par la courbure dans le processus de croissance. Nous travaillons en parallèle sur deux systèmes modèles dans l’oursin qui sont la régénération des épines chez l’adulte et la formation de la coquille chez le juvénile. Nos observations expérimentales montrent que le cytosquelette, et spécifiquement les fibres d’actine, pourrait jouer un rôle important dans cette interaction entre courbure et croissance. Actuellement, nous étudions cette hypothèse en développant un protocole pour caractériser l’organisation du cytosquelette. En parallèle nous travaillons aussi à un modèle de croissance qui couple l’énergie mécanique d’un réseau de fibre élastique à la géométrie du squelette auquel il s’attache.
Microscopie confocale à balayage, projection maximale. Calcium ajouté sur le stéréome pendant une nuit dans la coquille d’un juvénile en formation (gauche) et dans l’épine d’un adulte en régénération.
À lire aussi

Le youtubeur MonsieurLeChat au labo
Et si le caramel permettait de comprendre l’érosion d’un paysage ?Michaël Berhanu a reçu dans son laboratoire le youtubeur Monsieur Lechat, pour lui parler de dissolution. La vidéo est ici.
Face à la rigidification des sols, les racines développent une capacité d’auto-adaptation inédite
Une équipe du laboratoire Matière et Systèmes Complexes (MSC – Université Paris Cité/CNRS), impliquée dans une collaboration internationale, met en évidence un mécanisme original permettant aux racines de s’adapter à la rigidification des sols, un phénomène accentué...

Aquarium anéchoïque
Lorsque les parois d'une pièce réfléchissent trop le son, les réverbérations multiples peuvent vite rendre les conversations inintelligibles. Dans les salles de spectacle, les cinémas, et certaines salles de réunion, on cherche à traiter les surfaces, pour améliorer...

EvoDevMec2026 : Evolution, development and mechanics
We are happy to announce the forthcoming workshop entitled "Roadmap for EvoDevoMec", Nov. 2nd - 5th, 2026, Université Paris Cité. The link with evolutionary history of an organism is key to understand embryonic development. Much of the focus has been so far on genetic...