
Mécanique et comportements collectifs dans les tissus et amas cellulaires
Participants : Marc Durand, François Graner
Comment les cellules de notre corps s’assemblent, s’organisent entre elles, se déplacent, se déforment, et aboutissent à former des tissus biologiques vivants et fonctionnels ? Quoique sous le contrôle de l’information génétique, tous ces comportements sont du domaine de la physique, des forces et et mouvements, et font l’objet d’une riche recherche interdisciplinaire avec nos collègues biologistes, soit dans des systèmes en culture (“in vitro”), soit dans des organismes vivants (“in vivo”).
Analyse multi-échelles de tissus biologiques
Un matériau vivant est composé de cellules. Cependant, « vu de loin », le matériau vivant présente des comportements moyens, se traduisant par des lois de comportement proches de celles des milieux continus actifs. Pour comprendre la dynamique du vivant, il faut étudier les mécanismes physiques à l’origine de l’arrangement des cellules au sein d’un tissu biologique, et leur dynamique en cas d’écoulement, de croissance, de développement ou de différenciation. L’analyse multi-échelle permet de relier d’une part les processus cellulaires, et d’autre part les propriétés collectives au niveau du matériau tout entier. L’image suivante illustre les méthodes et les résultats de cette thématique. On y voit “l’écoulement” d’une monocouche cellulaire autour d’un obstacle circulaire (diamètre: 200 μm). L’étude de cet écoulement de Stokes nous renseigne sur la loi rhéologique constitutive du milieu cellulaire. Les résultats obtenus in vitro ont été intégrés à un effort de recherche à la fois in silico et in vivo. In silico par des simulations numériques multi-cellules, et des résolutions d’équations aux dérivées partielles, de manière à relier l’échelle de la cellule individuelle avec l’échelle des mouvements collectifs aboutissant à la morphogenèse du tissu. In vivo par la transposition des méthodes de mesure et d’analyse développées in vivo, à des situations concrètement observées lors de la morphogenèse de la mouche (notamment formation du dos, du cou, des organes sexuels mâles), ou dans le développement de l’embryon de souris ou d’oiseau.
Transition ordre-désordre dans les milieux cellulaires
L’organisation structurelle d’un tissu est essentielle à sa fonction biologique. Au cours des mouvements morphogénétiques, ont observe souvent le passage d’un état ordonné (pavage hexagonal où le cellules ont majoritairement 6 voisines) à un état désordonné (nombre important de cellules à 5 ou 7 voisines) du tissu. Ce comportement est très similaire à celui d’un cristal bidimensionnel soumis à une température croissante, suggérant ainsi que l’activité cellulaire agit comme une température effective. Nous avons caractérisé numériquement la transition ordre-désordre dans un milieu cellulaire fluide induite par l’agitation des interfaces. En particulier, nous avons mis en évidence l’existence d’une phase hexatique intermédiaire qui perd l’ordre translationnel mais conserve l’ordre orientationnel de la phase ordonnée, en accord avec le scénario de Kosterlitz, Thouless, Halperin, Nelson et Young (KTHNY) développé pour la fusion des solides bidimensionnels. Cet ordre hexatique intermédiaire a récemment (2022) été confirmé expérimentalement sur des monocouches cellulaires par une équipe de Leiden.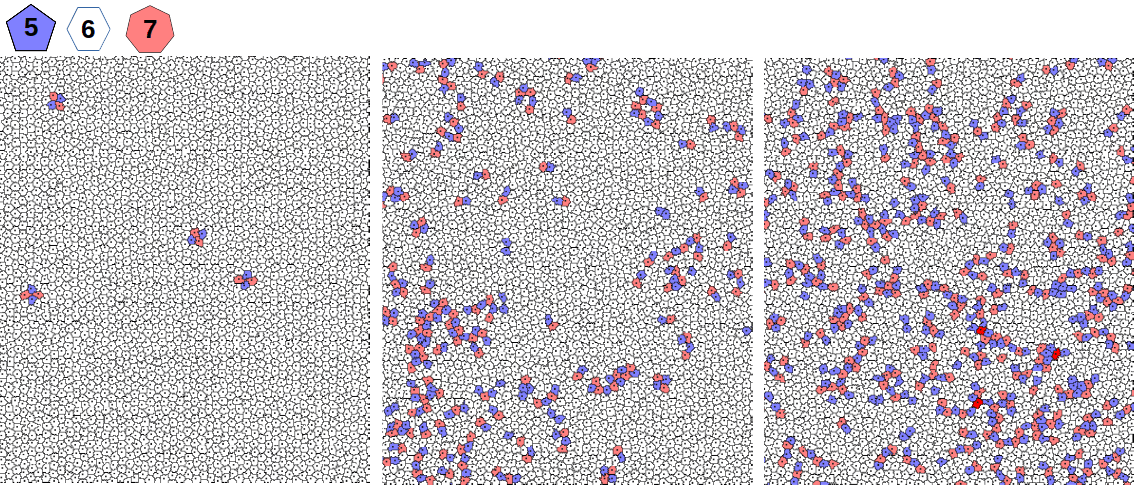
Cinétique de tri cellulaire induit par un contraste d’adhésion
La forme et la position des cellules biologiques évoluent durant la morphogenèse ou après une perturbation de leur configuration naturelle, comme dans le cas de la cicatrisation. In vitro, le tri spontané de deux types de cellules mélangées se produit dans des agrégats de cellules dissociées et donne des tissus homogènes et cohérents : un tel tri est une étape clé vers la reconstruction d’organes fonctionnels. Même des animaux adultes – comme l’hydre – peuvent se régénérer à partir d’un agrégat de cellules de différents types mélangées. Au cours des dernières décennies, il a été démontré expérimentalement que le contraste l’adhésion entre cellules homotypiques et hétérotypiques est le principal moteur du tri cellulaire dans les agrégats de cellules d’hydre. Mais il n’y avait pas consensus sur la cinétique de ce phénomène. Nous avons amélioré un modèle numérique permettant de caractériser cette cinétique sur des échelles de temps et d’espace inédites (jusqu’à 320 000 cellules évoluant sur 6 × 10^6 MCS). Cette étude a permis de montrer que l’évolution temporelle de la taille des agrégats de cellules homotypiques grossissaient selon une loi algébrique dont l’exposant était caractéristique d’une cinétique dominée par la coalescence de clusters arrondis, mettant ainsi fin au débat sur les résultats contradictoires obtenus dans des systèmes de plus petites tailles.

À lire aussi

Le youtubeur MonsieurLeChat au labo
Et si le caramel permettait de comprendre l’érosion d’un paysage ?Michaël Berhanu a reçu dans son laboratoire le youtubeur Monsieur Lechat, pour lui parler de dissolution. La vidéo est ici.
Face à la rigidification des sols, les racines développent une capacité d’auto-adaptation inédite
Une équipe du laboratoire Matière et Systèmes Complexes (MSC – Université Paris Cité/CNRS), impliquée dans une collaboration internationale, met en évidence un mécanisme original permettant aux racines de s’adapter à la rigidification des sols, un phénomène accentué...

Aquarium anéchoïque
Lorsque les parois d'une pièce réfléchissent trop le son, les réverbérations multiples peuvent vite rendre les conversations inintelligibles. Dans les salles de spectacle, les cinémas, et certaines salles de réunion, on cherche à traiter les surfaces, pour améliorer...

EvoDevMec2026 : Evolution, development and mechanics
We are happy to announce the forthcoming workshop entitled "Roadmap for EvoDevoMec", Nov. 2nd - 5th, 2026, Université Paris Cité. The link with evolutionary history of an organism is key to understand embryonic development. Much of the focus has been so far on genetic...